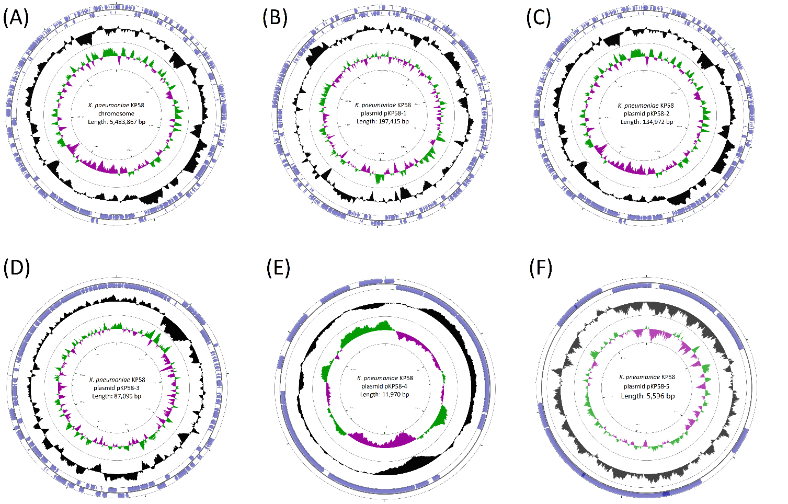
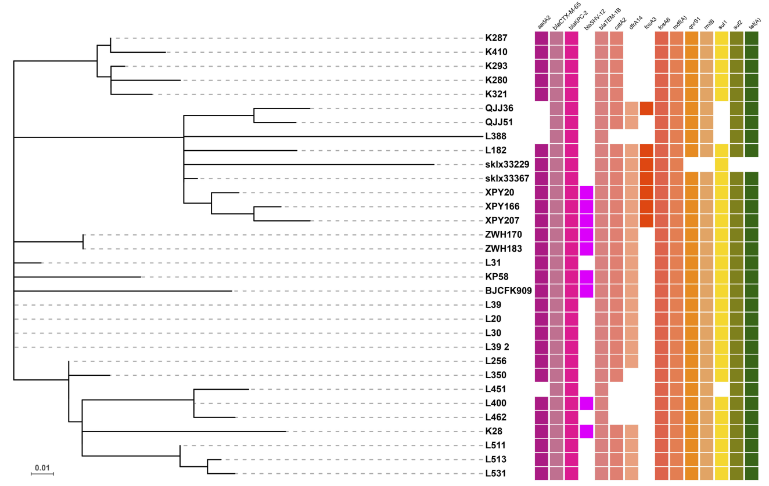
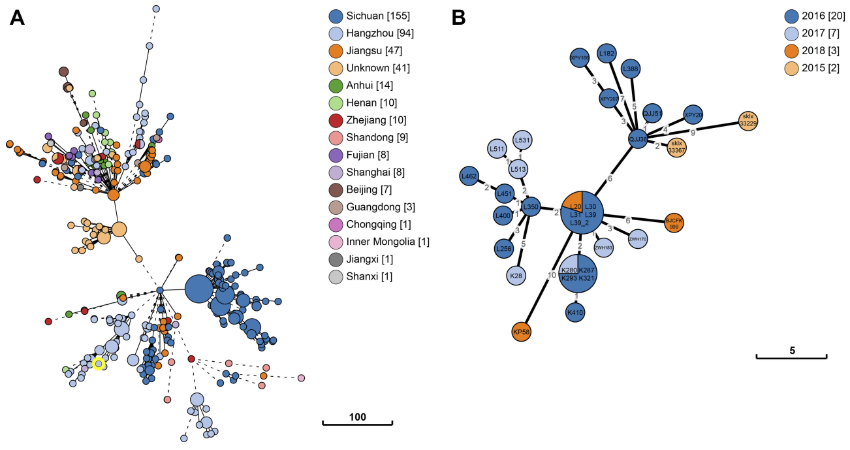
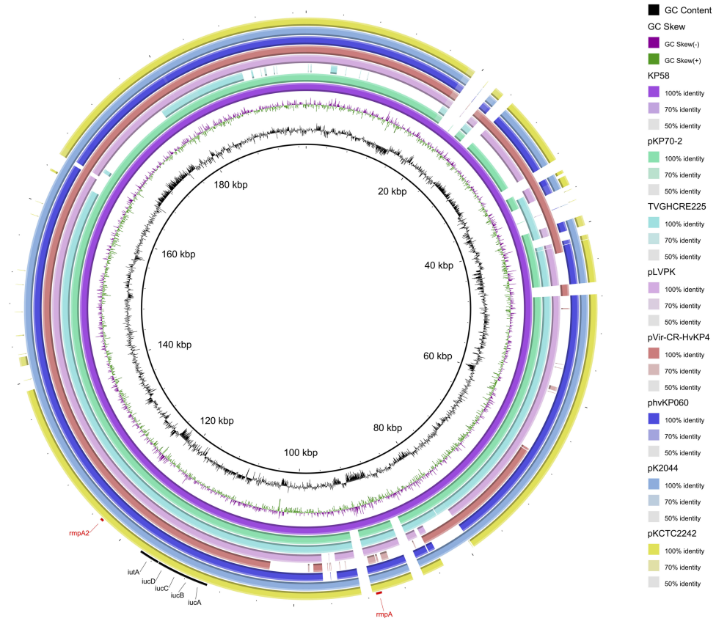
027-62435310 |
027-62435310 |
 service@benagen.com |
service@benagen.com |
- Home
- Services
-
Animal and plant genomes
-
Microbial genome
-
Transcriptional regulation
-
Microbial diversity
-
Animal and plant resequencing
-
Proteomics and metabolomics
-
Pure sequencing
-
Training course
-
- Medical
- Application
- Resources
- About us
- Contact us